محتوای آموزشیمحتوای آموزشی رشته تنوع زیستی
اندازهگیری تنوع زیستی فیلوژنتیک (تکامل نژادی) قسمت اول
درخت فیلوژنتیک
تنوع زیستی بعنوان بیولوژی تعداد و تفاوتها توصیف شده است. زیرا گونهها متفاوت از یکدیگرند ، معیارهای سنتی نظیر غنی بودن گونه یا یکنواختی، زمانی افزایش میابد که گونههای زیادی وجود دارد یا فراوانی به صورت کاملا یکسان بین گونهها توزیع شده است. نه تنها گونهها از یکدیگر متقاوت هستند، بلکه اندازه این تفاوتها بطور فوقالعادهای بسته به مجموعه گونههای مورد بحث تغییر میکند.
دو جزیره فرضی را درنظر بگیرید، که هریک فقط دو گونه از جانوران مهرهدار را با فراوانی یکسان دارند : جزیره اول دو پرنده از یک نوع و جزیره دوم، یک پرنده و یک پستاندار دارد. هر دو جزیره برای غنی بودن عدد 2 را دارند (برای مهرهداران) و مقدار ماکزیمم برای گونههای یکنواخت یکسان است. اگرچه شواهد به ما میگوید که یک پرنده بعلاوه یک پستاندار تنوع زیستی بیشتری را نسبت به دو پرنده ارائه میدهد. معیارهای تنوع فیلوژنتیک (تکامل نژادی) کمیت تفاوتها را معلوم میکند.
تفاوتهای میان گونهها میتواند با اندازهگیری تعدادی از ویژگیها شامل اندازه و شکل بدن، نیاز غذایی، تحمل فیزیولوژیکی در مقابل عوامل استرسزای مختلف و… مشخص گردد.
صفات و ویژگیهای خاصی ممکن است به دلایل مختلف مورد توجه ویژه یک محقق باشد. صفاتی از قبیل نقش فرض شده برای آنها در واسطهگری فعل و انفعالات میان گونههای مختلف (مانند اندازه منقار در پرندگان) یا اهمیت آنها در تحمل شرایط محیطی مختلف (مانند ضخامت برگ در گیاهان). اگرچه درجه تشابه یا تفاوت میان گونهها شدیدا بسته به انتخاب صفاتی است که اندازهگیری میشوند و بسیاری از صفات فقط قابل استفاده در گروههای خاص موجودات زنده میباشند (مانند نرخ فتوسنتز در گیاهان). یک روش کلی معمول برای اندازهگیری کمیت تنوع در میان گونهها، یک ارزیابی از روابط تکاملی گونهها در فرم طبقهبندی یا تکامل نژادی آنها میباشد. فیلوژنیهای مدرن از دادههای دنباله DNA مشتق میشوند که میتوانند برای همه موجودات زنده در درخت زندگی بدست آیند. فاصله تکامل نژادی بین دو گونه، یک تخمین زمانی از آخرین جد مشترک هر دو گونه است. به عبارت دیگر زمانی که هر یک از گونهها مستقل از دیگری تکامل یافته است. وقتی صفات مجزا همگرایی را نشان دهند، این نشاندهنده ارزشهای مشابه در حال تحول در اجداد مرتبط دور آنهاست، فاصله فیلوژنتیک یک نماینده برای نمایش بزرگی تفاوتهای فنوتیپی (در میان تعداد زیادی از صفات) که بین هر دو گونه انتظار میرود، ارائه میدهد.
زیستشناسان به دو دلیل اصلی علاقمند به اجزاء فیلوژنتیک تنوع زیستی هستند:
- برای ترکیب و آمیختن صریح گونههای متفاوت (از طریق یک ارزش مشترک کاربردی بین همه گونهها از باکتریها تا پستانداران) نه تنها تعدد گونهها بلکه الویتبندی حفاظت آنها.
- ثمر دادن بینشها نسبت به ساختار جوامع زیست محیطی
در مورد اول، درک اینکه تصمیمات سخت اولویتبندی مورد نیاز است تا در رابطه با سرمایهگذاری منابع محدود برای حفاظت گرفته شود، قبلا بحث شده است که هدف نباید فقط حفاظت از بزرگترین تعداد گونههای ممکن باشد بلکه حفاظت مجموعههایی از گونههاست که بیشترین تمایز در طبقهبندی را دارند یا بزرگترین انواع ممکن است از ویژگیهای بیولوژیکی را ارائه میدهند. برای این منظور، تلاشهای قابل توجهی در تعیین کمیت تمایز تکاملی (و در نتیجه ارزش حفاظت) گونههای مجزا یا تنوع فیلوژنتیک یک گروه از گونهها شده است.
از نظر تجزیه و تحلیل در زمینه بومشناسی اجتماعی، الحاق اطلاعات فیلوژنتیک تاریخچه نسبتا طولانی دارد. داروین (1859) فرض کرد که میان وابستگان (به عنوان مثال همنوعان) باید قویترین رقابت وجود داشته باشد. این امر تحقیقات بعدی را به سمت کشف نسبت گونه به تعداد جنسیت (یا جنسیت به خانواده) هدایت کرد که به طور بالقوه نشاندهنده نقش رقابت در ساختار جوامع زیستمحیطی است. اخیرا همین سوال مفهومی با استفاده از فیلوژنی مدرن مطرح شدهاست که شامل اطلاعات بیشتری در مورد روابط تکاملی نسبت به گروههای طبقهبندی میباشد. اگر واقعا وابستگان نزدیک قویترین رقابت را میان خود داشته باشند، جوامع محلی باید شامل گونههایی باشند که به صورت نسبتا دور بهم مرتبط هستند. متناوبا، عضویت گونهها در یک جامعه محلی ممکن است توسط تحمل شرایط محیطی فیزیکی (و نه بیولوژی) به بیشترین شکل ممکن محدود شود و اگر وابستگان نزدیک تحمل و بردباریهای مشابهی را به اشتراک گذارند، جوامع محلی باید شامل گونههایی باشند که بطور نسبتا نزدیکی به یکدیگر مرتبط هستند. برای آزمایش این فرضیهها محققان برخی از معیارهای تنوع فیلوژنتیک از پیشینه پژوهشی حفاظت را به کار گرفتهاند و همچنین برخی معیارهای اضافی متعلق به خودشان را نیز معرفی کردهاند. همانطوریکه در مسائل مربوط به زیستشناسی حفاظت و بومشناسی اجتماع مطرح میشود، تنوع فیلوژنتیک یک موضوع فوقالعاده مورد علاقه در پیشینه پژوهشی اخیر است. همینطور تعداد زیادی از معیارهای اندازهگیری تنوع فیلوژنتیک معرفی شدهاند و برای برخی از زیرمجموعه های این معیارها نیز آنالیزهایی برای ارزیابی فراوانی آنها یا درجهای که آنها تحت شرایط خاص در یک مجموعه یکدیگر را ملاقات خواهند کرد، انجام شدهاست. انتخاب معیارهای تنوع فیلوژنتیک در مطالعات تجربی کاملا تحت کنترل محققان است ولی با سه جنبه کلیدی یک سیستم خاص که بطور کامل (یا قسمتی از آنها) تحت کنترل محققان نمیباشند، متاثر خواهند شد: فرآیندهای زیرساختی، الگوهای مرتبط (جدای از تنوع فیلوژنتیک) و محدودیتهای داده.
انواع فرآیندهای تکاملی و زیستمحیطی روی مقادیر معیارهای تنوع فیلوژنتیک بصورت مستقیم یا غیر مستقیم اثر گذارند. فرآیندهای تکاملی که در دسته هایی فراتر از گونهها اتفاق می افتند (Macroevolutionary) الگوهایی ایجاد میکنند که موضوع یک بحث بزرگ برای ساختار جوامع فیلوژنتیک و انتخاب معیار میباشند. برای مثال حوزهای که در آن زایش گونه جدید زیستمحیطی است (مشتق شده از انتخاب واگرا) شکل فیلوژنی و حفاظت فیلوژنتیک از صفات مختلف را متاثر خواهد کرد. هر دوی اینها به نوبه خود روی حوزهای که همزیستی گونهها ممکن است توسط ارتباط درونی تحت تاثیر قرار گرفته باشد، اثرگذارند. فرآیندهای زیستمحیطی موثر بر ساختار جامعه از قبیل محدودیتهای محیطی روی سازگاری یا رقابت برای منابع نیز روی معیارهای تنوع فیلوژنتیک تاثیر خواهند داشت.
برخی الگوهای تجربی شامل توزیع تعداد گونهها و فراوانی میان سایتها و درجه تعادل در درخت فیلوژنتیک (شکل1a,b) ممکن است محدوده مقادیر ممکن که معیارهای مختلف میتوانند بگیرند را متاثر نمایند. همین فاکتور ممکن است روی درجهای که در آن یک معیار خاص از بقیه معیارها متمایز میگردد ، تاثیر داشته باشد. این الگوهای دیگر ممکن است توسط برخی از همان فرآیندهای تنوع فیلوژنتیک تحت تاثیر قرار گیرند اما میتوانند بطور مستقل و شاید روی اثر معیار تنوع فیلوژنتیک خودشان بدون توجه به فرآیندهایی که آنها را ایجاد کردهاند، تعیین کمیت شوند.
در پایان ممکن است محدودیتهای داده وجود داشته باشد. بطور مشخص طبیعت اطلاعات فیلوژنتیک ممکن است فقط اجازه نمایش توپولوژی یک درخت فیلوژنتیک یا فقط تخمین طول شاخههای متصل به گرهها را بدهد (شکل1a,b). بعلاوه ممکن است داده فقط به شرط حضور یا غیاب گونهها در مکانهای خاص بدون فراوانیهای نسبی در دسترس باشد. برخلاف فرآیندها و الگوهای ذکر شده تحت دو ملاحظات اول، این محدودیتها در تئوری در کنترل محققان هستند اما در عمل، مطالعات زیادی بر مبنای داده موجود انجام شده که ممکن است چنین محدودیتهایی را تحمیل کنند.
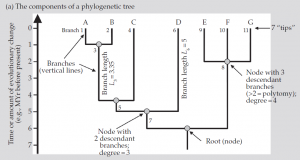
شکل (a1): اجزاء یک درخت فیلوژنتیک
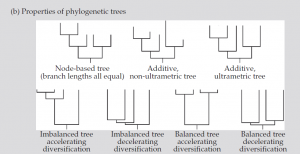
شکل (b1 ): اشکال و نمایشهای مختلف درخت که نشان دهنده خواص آنهاست
تعریف مشخصههای درختهای فیلوژنتیک:
یک درخت ریشهدار فیلوژنتیک ارتباطات تکاملی فرض شده میان گونهها یا سایر واحدهای زیستی از قبیل اجداد درون گونهها را خلاصه میکند. درختهای فیلوژنتیک میتوانند با استفاده از متدهای مختلفی تخمین زده شوند. برای اهداف این بحث فرض خواهیم کرد که برگهای درخت گونهها را نمایش میدهند (شکل1a,b). هر گره آخرین جد مشترک همه گونهها، از آن گره به پایین را ارائه می دهد (جائیکه شاخه ها جدا می شوند) و گره ریشه (که اغلب به آن ریشه گفته میشود) یک نقطه تکی است که از آن نقطه همه گونههای فرود آمده استنتاج می شوند، در نتیجه جهت زمانی کل درخت را میدهد. سادهترین نوع درخت فیلوژنتیک فقط توپولوژی را بدون هیچ اطلاعات دیگری روی طول شاخههای متصل به گرهها ارائه میدهد (طبقهبندی بر اساس دادههای مورفولوژیکی و برخی انواع دادههای مولکولی). ما به این درختها بعنوان درختهای مبتنی بر گره ارجاع خواهیم کرد.
یک شاخه در درخت فیلوژنتیک و طول شاخههای مربوط به آن ممکن است تجمع تغییرات تکاملی را نشان دهد که در این صورت برگها ممکن است در یک سطح قرار نگیرند چون نرخ تغییرات تکاملی در همه شاخهها یکسان نیست. در روش دیگر ، طولهای یک شاخه ممکن است مقیاسی برای نمایش گذر زمان در نظر گرفته شود بطوریکه تمام برگها در همان محل تنظیم و مرتب میشوند. هر دو نوع درخت فیلوژنیک افزایشی در نظر گرفته میشوند و نوع دوم علاوه بر افزایشی بودن، اولترامتریک نیز هست (همه فواصل از ریشه تا برگ به یک اندازه است). ما با طولهای کمی شاخهها به سراغ این درختها، بعنوان درختهای مبتنی بر فاصله، خواهیم رفت. گرهها معمولا به دو شاخه تقسیم میشوند و به همین صورت اجداد نیز به دو بخش تقسیم میگردند. Polytomies گره هایی هستند که در آنها اجداد به سه گروه یا بیشتر تقسیم می شوند.این وضعیت اغلب ناشی از محدودیت داده است. درجه یک گره تعداد شاخههای آنست، هر دو نوع شاخه متصل یعنی اجدادی و نوادگان شمرده میشوند (برای یک گره در درخت دودیی درجه گره 3 است). شکل درختان فیلوژنتیک میتوانند توسط دو خاصیت کلیدی مشخص شود: درجه تعادل آنها و درجهای که در آن حوادث واگرایی عمدتا در طی تکامل گروه دیر(واگرایی شتابدهنده) یا زود (واگرایی کندکننده) اتفاق افتاده است (واگرایی شتابدهنده توسط آمار γ مشخص میشود). در یک درخت متعادل کامل، همه برگها به فاصله تعداد گره ثابت از ریشه قرار دارند و این جمله معادل اینست که بگوییم که همه اجداد در زمان یکسانی از گره مربوطه خود منشعب شدهاند. در یک درخت کامل نامتعادل، یک جد نزولی از هر گره مستقیما به یک برگ وصل میشود بدون اینکه بیشتر دو شاخه شود. در یک درخت با γ پایین حوادث واگرایی در طی مراحل اولیه تکامل گروه متمرکز میشوند و بالعکس.
منبع و مطالعه بیشتر:
Vellend M, Cornwell WK, Magnuson-Ford K, Mooers A, Measuring phylogenetic biodiversity, biological diversity
مشاهده ادامه این مطلب فقط برای اعضای سایت فراگیر علمی تخصصی محیط زیست فراهم می باشد. خواهشمنداست جهت مشاهده کامل این نوشته، ثبت نام کنید و به حساب کابری خود وارد شوید. ثبت نام در اینجا کاملاً رایگان است.
مشاهده ادامه این مطلب فقط برای اعضای سایت فراگیر علمی تخصصی محیط زیست فراهم می باشد. خواهشمنداست جهت مشاهده کامل این نوشته، ثبت نام کنید و به حساب کابری خود وارد شوید. ثبت نام در اینجا کاملاً رایگان است.