اندازه گیری تنوع زیستی فیلوژنتیک (تکامل نژادی) بخش دوم
معرفی معیارهای تنوع فیلوژنتیک
تمامی مطالعات تجربی تنوع فیلوژنتیک با یک تخمین فیلوژنی برای گروهی از گونههای مورد علاقه آغاز میشود. محدوده این فیلوژنی تغییر میکند و ممکن است شامل همه گونههای شناخته شده در سراسر برخی مناطق گستردهتر باشد (مثلا همه پرندگان آمریکای جنوبی یا کل دنیا) یا فقط شامل آن گونههایی باشد که در نواحی خاص مورد مطالعه و بررسی، وجود دارند (مثل پرندگان یافت شده در 5 قطعه بررسی شده جنگلهای استوایی). برای راحتی کار، به این دو مورد به ترتیب با اسامی فیلوژنی منطقهای و فیلوژنی محلی رجوع میکنیم. بنابراین تعیین کمیت اندازه نسبی تنوع فیلوژنتیک در میان زیر مجموعههای کانونی گونهها که ممکن است بصورت رخدادهای همزمان در یک ناحیه محلی باشند (مثلا پرندگان در یک بخش جنگل استوایی)، یا یک مجموعه کاندیدا از گونهی پیشنهاد شده برای توجه حفاظت ویژه (گونههای لیست شده از پرندگان در معرض خطر در برزیل) مورد علاقه قرار گیرد. ما به بخشی از فیلوژنی ناحیهای یا محلی که شامل زیر مجموعههای کانونی گونهها میشود، به عنوان فیلوژنی زیر مجموعه ارجاع خواهیم کرد. دو نوع معیار مختلف کیفی تنوع فیلوژنتیک توسعه داده شدهاند. به معیار نوع اول به عنوان تنوعهای فیلوژنتیکی رجوع میشود که برای همه گونهها در یک فیلوژنی منطقهای یک امتیاز متمایز محاسبه مینمایند و سپس با محاسبه یکسری توابع (معمولا جمع) روی این امتیازها، برای زیرمجموعههای کانونی خاص این گونهها، منجر به ایجاد یک معیار برای تنوع فیلوژنتیک میگردد. معیارهای نوع 2 با یک فیلوژنی محلی (یا احتمالا یک فیلوژنی منطقهای)آغاز میگردند و برای یک زیر مجموعه کانونی از گونهها، آنها فقط به ویژگیهای فیلوژنی زیر مجموعه بستگی دارند. معیارهای نوع 1 بطور گستردهای در حفاظت محیط زیست استفاده شده است در حالیکه جوامع زیستمحیطی اکثرا معیارهای نوع 2 را بکار بردهاند، برخی در هر دو فیلد استفاده شدهاند. برای معیارهای نوع 1، برای اینکه اجازه دهد تا گونههای فردی بطریقی که وابسته به وضعیت سایر گونهها نباشند (مانند اینکه آیا تا بحال حمایت شدهاند یا نه)، رتبه بندی شوند، انگیزه پشت اولین محاسبه امتیازات گونههای فردی را ثابت کرد به جای اینکه بطور موثر اجازه وابستگی آنها به مجموعه گونههای کانونی را بدهد (مانند معیارهای نوع 2).
معیارهای نوع 1 تنوع فیلوژنتیک در دو مرحله محاسبه میشوند. ابتدا یک اندیس تمایز برای هر گونه محاسبه میشود و سپس این مقادیر وارد یک تابع جداگانه برای خلاصه کردن امتیازات هر مجموعه کانونی از گونهها میشوند. در مرحله یک ، پنج اندیس متفاوت تمایز گونهها در پیشینه پژوهشی استفاده میشود: تمایز طبقهبندی(TD)، اصالت گونهها (SO)، شاخههای خروجی (PE)، تاریخچه تکاملی گونه (SEH) و اصالت گونهها در یک مجموعه (OSS) (جدول1 را ببینید.). دو اندیس اول (SO,TD) فقط بر اساس درختهای فیلوژنتیک مبتنی بر گره هستند و سه اندیس بعدی (PE,SHE,OSS) بر اساس درختهای مبتنی بر فاصله میباشند. در مرحله دوم بیشتر توابع جمع هستند که بطور واضح قصد یکی کردن غنای گونهها درون یک معیار را دارند (وقتی سایر موارد برابر هستند، گونههای بیشتر تنوع فیلوژنتیکی بیشتر را ارائه میکنند). با فراوانی داده گونهها، بکار گرفتن یک روش مشابه رقیق کردن برای ایجاد یک اندیسی که مجموع مورد انتظار در یک نمونه از Xهای تکی انتخاب شده بصورت تصادفی از یک اجتماع را منعکس میکند هم امکانپذیر است. اما این روش بندرت در ادبیات استفاده میشود. در تئوری نیز امکان محاسبه متوسط مقادیر تمایز وجود دارد اگرچه این نیز به ندرت استفاده میشود.
در نگاه اول، تعداد معیارهای مختلف نوع 2 خیلی بزرگ بنظر میآید اما تمایز میان بسیاری از آنها فقط بر اساس طبیعت داده ورودی است بجای اینکه بر اساس معادلهای باشد که توسط آن داده وارد میشود. دو ویژگی داده بطور خاص اجازه میدهد مجموعههای گوناگونی از معیارها با هم یکی شوند. ابتدا طبیعت درخت فیلوژنتیک است (شکل 1). اگر فقط توپولوژی درخت تخمین زده شده باشد، فواصل داخل درخت بسادگی توسط شمردن گرهها در امتداد مسیر دلخواه محاسبه میگردد (بین دو برگ). طولهای کمی شاخهها اجازه میدهند که فواصل یک محدوده پیوسته از مقادیر را بگیرند. معیارهای مبتنی بر شمارش گرهها بطور موثری موارد خاصی از معیارهای مبتنی بر فاصله هستند که در همه آنها طول همه شاخهها یک در نظر گرفته میشود. ثانیا نیز طبعیت دادههای تکراری گونهها میباشد. همچنین ممکن است تخمینهای کمی از تکرارهای گونهها یا فقط حضور و غیاب آنها باشد. معیارهای دسته دوم موارد خاصی از معیارهای قبلی هستند که در هر گونه همان فراوانی را دارند. چهار نوع متمایز از معیارهای نوع 2 پیشنهاد شده است(توصیف شده در جدول 2). هریک از آنها می توانند فراوانی داده را در صورت دردسترس بودن یکی کنند و همچنین ممکن است به عنوان انحراف از مقادیر مورد انتظار بر اساس مدلهای تهی بیان شوند.
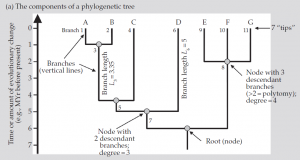
شکل(a) :1 اجزاء یک درخت فیلوژنتیک
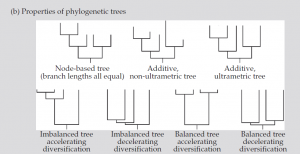
شکل (b):1 اشکال و نمایشهای مختلف درخت که نشان دهنده خواص آنهاست
جدول 1 اندیسهای تمایز گونه برای استفاده در معیارهای نوع 1 تنوع فیلوژنتیک
| اندیس | توضیح | مرجع |
| تمایز طبقهبندی (TD) |
تعداد گره موجود بین گونه و ریشه درخت (استاندارد شده توسط تقسیم بر مجموع این امتیازات در سراسر گونه و ضرب در 100) اصلاح: جهت انجام شمارش برای polytomieها، تعداد فرزندان را به جای تعداد گرهها بشمارید. |
Vane-Wright et al. (1991)
May (1990) |
| اصالت گونه (SO) |
اگر یک گره دارای تعداد گونه خروجی بیشتری نسبت به گره خواهر خود است،به آن گره در درخت مقدار 1 نسبت دهید و در غیر اینصورت مقدار 0. مجموع مقادیر در گرههای بین یک گونه و ریشه.، مقادیر کوچکتر تمایز بزرگتر را مشخص میکنند. اصلاح: همانند بالا، اما به هر گره یک مقدار معادل با تعداد گونههایی که از آن گره خارج می شوند ، اختصاص دهید. به عنوان اصالت گونه وزندار به آن ارجاع میشود(WSO). |
Nixon & Wheeler (1992)
|
| لبه خروجی (PE) | طول هر شاخه متصل به گونه تا بقیه درخت ناحیهای | Altschul & Lipman (1990) |
| تاریخچه تکامل گونه (SEH) |
بخشی از یک درخت فیلوژنتیک مربوط به یک گونه.شاخههای مشترک بصورت یکسان بین اجداد نوادگان تقسیم میشوند (انشعابات برابر)، برای مثال در یک درخت بدون polytomieها،بخشی از شاخه که به گونهای تخصیص یافته که n گره دورتر ازآن شاخه است،برابر است. اصلاح : همانند بالا، اما شاخههای مشترک بطور مساوی بین نوادگان گونه تقسیم میشوند (تقسیم عادلانه یا SEH_fair). رجوع شود به تمایز تکامل گونه |
Redding & Mooers (2006)
Redding et al. (2008) |
| اصالت گونه در یک مجموعه (OSS) |
مقادیر برای هر گونه که “به حداکثر رساندن عدم تشابه مورد انتظار بین دو گونه تصادفی انتخاب شده از مجموعه”. راه ساده تری برای توصیف این معیار وجود ندارد. |
Pavoine et al. (2005b) |
مدل خلاصه شده با حروف بزرگ در متن استفاده میشود.معادلات نشان داده نشدهاند چون دربیشتر موارد یا توضیحات شفاهی خیلی ساده هستند (TD,PE) یا نوشتن یک معادلهای که معنای بیشتری از معیار را روشن میسازد، امکانپذیر نیست (SO,OSS).
مدلهای تهی:
هریک از معیارهایی که تاکنون توصیف کردهایم میتوانند به فرم خام خود یا بعنوان انحرافهایی از دادههای مشتق شده مورد انتظار از یک مدل تهی توضیح داده شوند. در عمل، مدلهای تهی در ادبیات حفاظت بکار گرفته نشدهاند اما در ادبیات جامعه اکولوژی رایج هستند. بعلاوه مدلهای تهی اکثرا همیشه به همراه دادههای حضور-غیاب در کوچکترین مقیاسها استفاده میشوند. هدف اصلی در ساخت یک مدل تهی اینست که بپرسیم چه توزیعی از مقادیر برای مجموعههای کانونی گونههایی که در آنها هیچ ساختار فیلوژنتیکی وجود ندارد ولی در همه ویژگیهای دیگر یکسان هستند (از قبیل غنای گونه) ، مورد انتظار است.
دو کلاس اصلی از مدلهای تهی استفاده شدهاند. ابتدا برای یک مجموعه کانونی از گونههای S، یکی از کلاسها میتواند نمونههای تصادفی تکراری از گونههای S را که بطور تصادفی از درخت فیلوژنتیک منطقهای انتخاب شدهاند را دریافت نماید. در اصل این کار به مثابه مخلوط کردن هویت گونههای یکسان بطور تصادفی در میان برگهای یک درخت فیلوژنتیک میباشد. به روش دیگر، با یک ماتریس داده گونهها -به تفکیک- سایت، یک کلاس میتواند مکررا حضور گونهها در میان سایتها را مخلوط نماید تا اساسا بصورت تصادفی معین کند که کدام گونهها همزمان اتفاق میافتند در حالتی که تکرار گونهها در سراسر سایت ثابت نگه داشته شده است. همچنین ممکن است محدودیتهای اضافی متنوعی در رویه مخلوط کردن قرار داده شود مانند همزمان حفظ کردن غنای گونههای هر سایت و تکرار هر یک از گونهها به محض اینکه یک مدل تهی ساخته شد ، معیارهای مورد علاقه برای هر مجموعه شبیهسازی شده گونهها محاسبه میگردند و مقادیر مشاهده شده میتواند یا به عنوان تعداد استاندارد انحراف از میانگین مورد انتظار و یا احتمال بدست آوردن یک مقدار کمتر (یا بیشتر) از مقدار مشاهده شده منحصرا اتفاقی، بیان شوند. از آنجائیکه که در بسیاری موارد، توزیع مقادیر به غنای گونهها بستگی دارد، معیارهای توضیح داده شده در این روش معمولا تبدیلهای یکنواخت از مقادیر اولیه نیستند. برای مثال تنوع فیلوژنتیک (PD) ممکن است در دو جامعه مختلف با تعداد گونههای متفاوت، معادل باشد اما آنکه گونههای کمتری دارد، استثناءهای تهی کمتری خواهد داشت و بنابراین مقادیر دوباره بیان شده در جامعهای که گونههای کمتری دارد، بالاتر خواهد بود.
جدول 2- معیارهای نوع 2 تنوع فیلوژنتیک
| معیار | نسخه حضور-غیاب(PA) | نسخه تکرار-وزن (AW) | معادله | مراجع |
| تنوع فیلوژنتیک (PD) |
مجموع طول همه شاخهها در بخشی از درخت فیلوژنتیک که مربوط به مجموعه کانونی گونه میباشد (PD,PDn) |
برای درخت زیر مجموعه ، حاصلضرب تعداد شاخهها در وزن متوسط طول شاخه ، با وزنهای مساوی در میانگین تکرار گونههایی که در آن شاخه مشترک هستند*(Pdaw,PDnaw) |
PA: Faith (1992)
AW: Barker (2002) |
|
| متوسط فاصله فیلوژنتیک (MPD)
|
متوسط فاصله فیلوژنتیک بین دو زوج از گونه در یک مجموعه کانونی (MPD,MPDn)
|
متوسط فاصله فیلوژنتیک بین زوجهای غیرتکراری (یا سایر واحدهای تکرار شونده)به جز همان جفت گونه (MPDaw,MPDnaw) اصلاح : متوسط فاصله فیلوژنتیک بین زوجهای غیرتکراری (یا سایر واحدهای تکرار شونده )شامل همان جفت گونه |
PA: Webb (2000)
AW:Warwick& Clarke (1995 |
|
| مجموع فواصل فیلوژنتیک
(SPD) |
مجموع فواصل فیلوژنتیک بین دو زوج از گونه که برابر حاصلضرب MPD در تعداد جفت گونهها است |
حاصلضرب فراوانی-وزن در تعداد جفت گونهها. |
Rao (1982); Warwick& Clarke(1995) | |
| متوسط فاصله نزدیکترین همسایه (MMND) | متوسط فاصله فیلوژنتیک از هر گونه تا نزدیکترین وابستگانش در مجموعه کانونی گونه (MNND,MNNDn) | متوسط وزنی فاصله فیلوژنتیک از هر گونه تا نزدیکترین بستگانش ، با وزنهای مساوی تکرار گونه (MNNDaw,MNNDnaw) | PA: Webb (2000)
AW: none |
فرمهای خلاصه شده مربوط به برچسبها در قسمت PCA (شکل 1 را ببینید) ؛ معیار مبتنی بر گره=n ،B تعداد شاخهها در درخت، Li طول هر شاخه i ،Ai میانگین فراوانی (اندازهگیری شده در هر واحد)گونههایی که در شاخه i مشترک هستند، dmn فاصله فیلوژنتیک بین گونهm, n ، am فراوانی گونه m(با داده حضور و غیاب همه گونهها همان فراوانی را دارد)؛ S تعداد گونهها در مجموعه کانونی، aw معیار فراوانی-وزن.
*این تفسیر ما از اینست که چطور متد تکراری Baker (برای درختهای بدون ریشه)ممکن است به سادگی برای یک درخت ریشه دار استفاده شود.
+ برای درختهای اولترامتریک ، نسخه حضور و غیاب MPD معادل دوبرابر معیار تنوع گونههای فیلوژنتیک (PSV) میباشد که از واریانس مورد انتظار در میان گونهها در یک صفت بیطرفانه تکامل یافته، مشتق میشود. معیار یکنواختی گونه فیلوژنتیک (PSE) نسخه مجددا مقیاسبندی شده فراوانی-وزن MPD میباشد(هر دو معیار PSV و PSE با استفاده از درخت فیلوژنتیک با طول شاخه مقیاسبندی شده محاسبه شدهاند و همه فواصل از ریشه تا برگ 1 هستند.).
++ Helmus et al. (2007) غنای گونه فیلوژنتیک (PSR) را به جای تعداد جفت گونه ها، با ضرب MPD در تعداد گونه ها محاسبه می کند؛ اگرچه این دو گزینه بصورت یکنواخت با هم مقیاس می شوند.
معیارهای مختلف میتوانند مقایسه گردند و از لحاظ کمی و کیفی ارزیابی شوند. از نظر مقایسههای کیفی، یک انتخاب محقق از معیارها، ممکن است به اینکه میخواهند چه اطلاعاتی در معیار منعکس شود (مثلا وابستگان نزدیک گونهها یا مجموعه کامل روابط آنها) بستگی داشته باشد به جای اینکه به هر معیار کمی از پیش تعیین شده وابسته باشد (مثلا حساسیت به ترکیب یک انجمن غیر تصادفی). هر دو ملاحظات کمی و کیفی در بخش اطلاع نامه مورد بحث قرار میگیرند و برای انجام مقایسههای کمی، ما یک مجموعه از آنالیزهای شبیهسازی شده را انجام دادیم. هدف این شبیهسازیها پاسخگویی به سه سوال زیر است: (1) کدام معیارها نسبت به یکدیگر فراوانی دارند؟ (2) چگونه معیارها با توجه به حساسیت آنها به فرآیندهای ترکیب جوامع غیر تصادفی مختلف ، با هم مقایسه میشوند و (3) چگونه شکل درخت روی پاسخ به سوالات (1)و (2) تاثیرگذار خواهد بود.
تنوع فیلوژنتیک در حفاظت:
به حدی اختصاص امتیاز تمایز گونههای فردی ارزشی پر اهمیت در زیستشناسی حفاظت است که پژوهشگران و محققان باید به پیدا کردن معیارهای مفید نوع 1 ادامه دهند. مجموعها در همه گونهها برای همه معیارهای نوع 1 نسبتا با شدت بالا با یکدیگر و PD همبستگی دارند. درختهای مبتنی بر فاصله بطور کامل تری نسبت به درختهای مبتنی بر گره ، ارتباطات تکاملی را نشان میدهند و چون استفاده از معیارهای مبتنی بر گره بنظر میرسد تا حد زیادی توسط ملاحظات لجستیک برانگیخته شوند (فقدان درخت مبتنی بر فاصله) ، معیارهای مبتنی بر فاصله ، به وضوح قابل ترجیح بنظر میرسند. در میان معیارهای مبتنی بر گره و مبتنی بر فاصله فراوانی کاملا بالاست (برای همه انواع درخت r>0.9 یعنی همبستگی بالای 0.9). به این ترتیب، یک ملاک مناسب برای انتخاب میان معیارها ، سادگی مفهومی و ریاضی آنهاست. برای هر معیار مبتنی بر گره ، ما معیار تمایز طبقهبندی (TD) را بدلیل سادهتر بودن و سر راست تر بودن از هر دو نسخه معیارهای اصالت گونهها (OS،WOS، جدول 1) پیشنهاد میدهیم. برای معیارهای مبتنی بر فاصله ، معیار اصالت گونهها در یک مجموعه (OSS)بطور مفهومی سادگی کمتر دارد ، نیاز به یک درخت اولترامتریک دارد ، از لحاظ ریاضی بسیار پیچیدهتر است و از نسبت به دیگران تعداد PD کمتری تسخیر میکند، بنابراین حتی با وجود این حقیقت که همبستگی پایینتری با سایر معیارها را نشان میدهد ، ما هنوز استفاده از آن را پیشنهاد نمیکنیم. معیارهای لبههای خروجی (PE) و تاریخچه تکامل گونهها (SEH) هر دو از از لحاظ مفهومی و ریاضی ساده هستند (PE بسیار سادهتر است). اگرچهPE و SEH تا حد زیادی روی بیشتر اشکال درختها فراوانی دارند،SEH شامل اطلاعات بیشتری است و برای مثال، به عنوان تمایز تکاملی هر یک از گونهها در یک جفت از بستگان نزدیک را ، وقتی که جفت گونه خودش یک تمایز تکاملی است، شناسایی خواهد کرد در حالیکه PE در این مورد کاملا کوچک خواهد بود. مهم نیست که کدام روش تخصیص شاخههای به اشتراک گذاشته شده در محاسبه SEH بکار میرود (برای همه انواع درخت r≥0.98).
در برخی موارد ممکن است یک زیستشناس حفاظت علاقهای به اولویتبندی گونهها نداشته باشد، اما علاقمند به اولویتبندی سایتها براساس گونههایی که شامل شدهاند، یا در درک چگونگی فرآیندهای اکوسیستم بسته به تنوع فیلوژنتیک ، به جای فقط غنا، از گونههای یک جامعه باشد. در چنین مواردی نیازی به تخصیص امتیازات انفرادی نیست و مناسبتر است که PD بعنوان یک پروکسی سادهتر برای مقدار تاریخچه تکامل و بنابراین تغییرات صفت در یک جامعه استفاده شود.
تنوع فیلوژنتیک در محیط زیست جامعه:
وقتی اکولوژیستهای جامعه علاقمند به استفاده از اطلاعات فیلوژنتیک برای ارزیابی درجهای که در آن ترکیب جامعه با توجه به صفات گونهها یا ارتباط آنها غیرتصادفی بوده است ، مناسبترین راه استفاده از مدل تهی نسخه اصلاح شده معیارهای نوع 2 میباشد. متوسط مقدار برخی معیارها (در فرم خام آنها) با غنای گونه همبستگی دارد (MNND گونههایی که به یک جامعه اضافه شدهاند را کاهش میدهد.) و حتی برای آنهایی که (مثلا MPD) در میان جوامع ترکیب شده بصورت تصادفی متمایز نیستند، ممکن است همبستگی خوبی با غنای گونه وجود داشته باشد. محدوده مقادیر MPD ، برای مثال، وقتی تعداد گونهها افزایش مییابد، کاهش پیدا میکند. به این ترتیب، میزانی که اعضای یک جامعه در انتخاب غیرتصادفی از فیلوژنی نشان میدهند، بطور کلی نمیتواند با استفاده از مقادیر خام اولیه برای معیارهای نوع 2 ارزیابی شود. در شرایط مدلهای تهی، مخلوط کردن گونهها در میان برگهای فیلوژنی، بنظر سادهترین راه است چون بطور مستقیمتری ویژگیهای کلیدی اطلاعات مورد علاقه را بصورت تصادفی توزیع میکند.
نکات کلیدی:
- معیارهای تنوع فیلوژنتیک در بیولوژی حفاظت استفاده میشوند که در آن مطلوبست که ابتدا امتیاز تمایز گونه تکی تخصیص یابد و همچنین در اکولوژی جامعهای بکار میروند که در آن علاقه به ارزیابی درجهای وجود دارد که در آن درجه گونهها در یک جامعه، یک زیر مجموعه غیرتصادفی از گونهها در یک درخت فیلوژنتیک را نشان میدهند.
- معیارهای بکار رفته در بیولوژی حفاظت درجه بالایی از فراوانی (برای مثال همبستگی قوی) را بین یکدیگر و با تنوع فیلوژنتیک کلی نشان میدهند(مجموع طول شاخههای متصل به گونه در یک مجموعه کانونی). بریا یک درخت فیلوژنتیک مبتنی بر گره (که بعنوان مثال هیچ اطلاعاتی روی طول شاخهها وجود ندارد) ، معیار تنوع طبقهبندی (TD)بطور مفهومی سادهترین است و برای درختان مبتنی بر فاصله معیارهای لبه خروجی(PE) و تاریخچه تکامل گونه (SEH)هر دو از لحاظ مفهومی و ریاضی سادهترین هستند.
- در آزمایش ترکیب جامعه غیر تصادفی، نیاز است که معیارها بر اساس مدلهای تهی استاندارد شوند تا بتوانند وابستگی ذاتی به غنای گونه را حذف نمایند. برای چنین معیارهای اصلاح نشده، آنهایی که از درختهای مبتنی بر گره محاسبه میشوند ، میتوانند بطور جدی گمراه کننده باشند. فراوانی در میان آنهایی که از روی درختهای مبتنی بر فاصله محاسبه شدهاند ، بطرز زیادی بالاست. ما استفاده از درختهای مبتنی بر گره در محیط زیست جامعه فیلوژنتیک را پیشنهاد نمیکنیم.
- حساسیت معیارهای تنوع فیلوژنتیک بشدت بسته به شکل درخت فیلوژنتیک میباشد، با فیلوژنتیک خوشهای برخی موارد بیشتر قابل شناسایی هستند (مانند درختهای متعادل) و با فیلوژنتیک فوق پراکنده برخی دیگر بیشتر شناسایی میشوند (درختهای نامتعادل با شتاب تنوعسازی مثبت). برای محققان تجربی به حساب آوردن شکل درخت خاص خودشان در قدرت آماری مهم است.
منبع:
Vellend M, Cornwell WK, Magnuson-Ford K, Mooers A, Measuring phylogenetic biodiversity, biological diversity (chapter 14).
مطالب مرتبط با تنوع فیلوژنتیک:
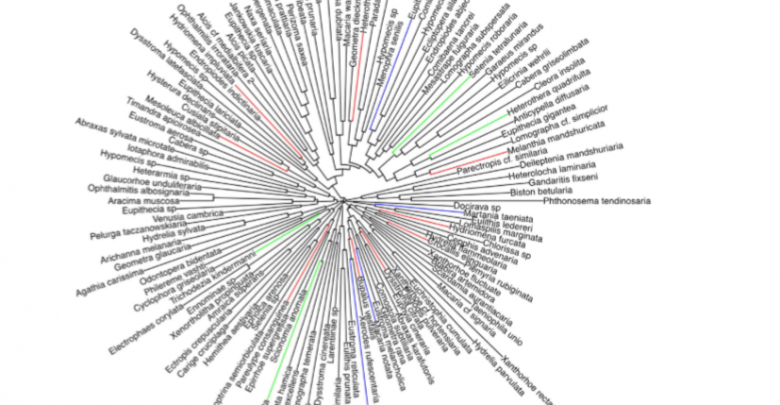
Zou Y, Sang W, Hausmann A, Axmacher J, (2016), High phylogenetic diversity is preserved in species-poor high-elevation temperate moth assemblages, Scientific Reports | 6:23045 | DOI: 10.1038/srep23045
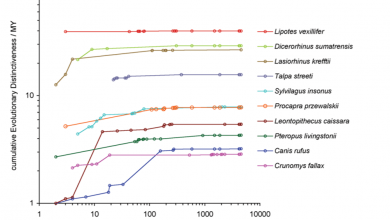
Isaac NJB, Turvey ST, Collen B, Waterman C, Baillie JEM (2007) Mammals on the EDGE: Conservation Priorities Based on Threat and Phylogeny. PLoS ONE 2(3): e296. doi:10.1371/journal.pone.0000296
مشاهده ادامه این مطلب فقط برای اعضای سایت فراگیر علمی تخصصی محیط زیست فراهم می باشد. خواهشمنداست جهت مشاهده کامل این نوشته، ثبت نام کنید و به حساب کابری خود وارد شوید. ثبت نام در اینجا کاملاً رایگان است.
مشاهده ادامه این مطلب فقط برای اعضای سایت فراگیر علمی تخصصی محیط زیست فراهم می باشد. خواهشمنداست جهت مشاهده کامل این نوشته، ثبت نام کنید و به حساب کابری خود وارد شوید. ثبت نام در اینجا کاملاً رایگان است.